 W
WAn antorbital fenestra is an opening in the skull that is in front of the eye sockets. This skull character is largely associated with archosauriforms, first appearing during the Triassic Period. Among extant archosaurs, birds still possess antorbital fenestrae, whereas crocodylians have lost them. The loss in crocodylians is believed to be related to the structural needs of their skulls for the bite force and feeding behaviours that they employ. In some archosaur species, the opening has closed but its location is still marked by a depression, or fossa, on the surface of the skull called the antorbital fossa.
 W
WAn arctometatarsalian organism is one in which the proximal part of the middle metatarsal is pinched between the surrounding metatarsals. The trait appears to be highly homoplastic, common in certain sorts of dinosaurs accustomed to running, to evenly transmit force to the metatarsals.
 W
WArmour or armor in animals is external or superficial protection against attack by predators, formed as part of the body, usually through the hardening of body tissues, outgrowths or secretions. It has therefore mostly developed in 'prey' species.
 W
WIn zoology, a club is a bony mass at the end of the tail of some dinosaurs and of some mammals, most notably the ankylosaurids and the glyptodonts, as well as meiolaniid turtles. It is thought that this was a form of defensive armour or weapon that was used to defend against predators, much in the same way as a thagomizer, possessed by stegosaurids, though at least in glyptodonts it is hypothesized it was used in fighting for mating rights. Among dinosaurs, the club was present mainly in ankylosaurids, although the sauropod Shunosaurus also possessed a tail club. The tail club is most often depicted on Ankylosaurus, especially in encounters with larger predators such as Tyrannosaurus. Victoria Arbour has established that ankylosaurid tails could generate enough force to break bone during impacts.
 W
WA crop is a thin-walled expanded portion of the alimentary tract used for the storage of food prior to digestion. This anatomical structure is found in a wide variety of animals. It has been found in birds, and in invertebrate animals including gastropods, earthworms, leeches, and insects.
 W
WDinosaur teeth have been studied since 1822 when Mary Ann Mantell (1795-1869) and her husband Dr Gideon Algernon Mantell (1790-1852) discovered an Iguanodon tooth in Sussex in England. Unlike mammal teeth, individual dinosaur teeth are generally not considered by paleontologists to be diagnostic to the genus or species level for unknown taxa, due morphological convergence and variability between teeth. and many historically named tooth taxa like Paronychodon and Richardoestesia are today considered nomina dubia, and are used as form taxa to refer to isolated teeth from other localities displaced considerably in time and space from the type specimens. However, it is possible to refer isolated teeth to known taxa provided that the tooth morphology is known and the teeth originate from a similar time and place.
 W
WEpipophyses are bony projections of the cervical vertebrae found in archosauromorphs, particularly dinosaurs. These paired processes sit above the postzygapophyses on the rear of the vertebral neural arch. Their morphology is variable and ranges from small, simple, hill-like elevations to large, complex, winglike projections. Epipophyses provided large attachment areas for several neck muscles; large epipophyses are therefore indicative of a strong neck musculature.
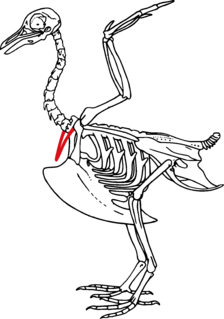 W
WThe furcula or wishbone is a forked bone found in birds and some other species of dinosaurs, and is formed by the fusion of the two clavicles. In birds, its primary function is in the strengthening of the thoracic skeleton to withstand the rigors of flight.
 W
WThe gizzard, also referred to as the ventriculus, gastric mill, and gigerium, is an organ found in the digestive tract of some animals, including archosaurs, earthworms, some gastropods, some fish, and some crustaceans. This specialized stomach constructed of thick muscular walls is used for grinding up food, often aided by particles of stone or grit. In certain insects and molluscs, the gizzard features chitinous plates or teeth.
 W
WA horn is a permanent pointed projection on the head of various animals that consists of a covering of keratin and other proteins surrounding a core of live bone. Horns are distinct from antlers, which are not permanent. In mammals, true horns are found mainly among the ruminant artiodactyls, in the families Antilocapridae (pronghorn) and Bovidae. Cattle horns arise from subcutaneous connective tissue and later fuse to the underlying frontal bone.
WAn infratemporal fenestra, also called the lateral temporal fenestra or simply temporal fenestra, is an opening in the skull behind the orbit in some animals. An opening in front of the eye sockets, conversely, is called an antorbital fenestra. Both of these openings reduced the weight of the skull. Infratemporal fenestrae are commonly seen in the fossilized skulls of dinosaurs.
 W
WThe manus is the zoological term for the distal portion of the fore limb of an animal. In tetrapods, it is the part of the pentadactyl limb that includes the metacarpals and digits (phalanges). During evolution, it has taken many forms and served a variety of functions. It can be represented by the hand of primates, the lower front limb of hoofed animals or the fore paw and is represented in the wing of birds, bats and prehistoric flying reptiles (pterosaurs), the flipper of marine mammals and the 'paddle' of extinct marine reptiles, such as plesiosaurs and ichthyosaurs.
 W
WA neck frill is the relatively extensive margin seen on the back of the heads of reptiles with either a bony support such as those present on the skulls of dinosaurs of the suborder Marginocephalia or a cartilaginous one as in the frill-necked lizard.
 W
WA neural spine sail is a large, flattish protrusion from the back of an animal colinear with the spine. Many extinct species of amphibians and amniotes have extended neural spines growing from their back vertebrae. These are thought to have supported a sail. Paleontologists have proposed a number of ways in which the sail could have functioned in life.
 W
WOsteoderms are bony deposits forming scales, plates or other structures based in the dermis. Osteoderms are found in many groups of extant and extinct reptiles and amphibians, including lizards, crocodilians, frogs, temnospondyls, various groups of dinosaurs, phytosaurs, aetosaurs, placodonts, and hupehsuchians.
 W
WThe palpebral bone is a small dermal bone found in the region of the eye socket in a variety of animals, including crocodilians and ornithischian dinosaurs. It is also known as the adlacrimal or supraorbital, although the latter term may not be confused with the supraorbital in osteichthyan fishes. In ornithischians, the palpebral can form a prong that projects from the front upper corner of the orbit. It is large in heterodontosaurids, basal ornithopods such as Thescelosaurus and Dryosaurus, and basal ceratopsians such as Archaeoceratops; in these animals, the prong is elongate and would have stuck out and over the eye like a bony eyebrow. As paleoartist Gregory S. Paul has noted, elongate palpebrals would have given their owners fierce-looking "eagle eyes". In such cases, the expanded palpebral may have functioned to shade the eye.
 W
WThe pes is the zoological term for the distal portion of the hind limb of tetrapod animals. It is the part of the pentadactyl limb that includes the metatarsals and digits (phalanges). During evolution, it has taken many forms and served a variety of functions. It can be represented by the foot of primates, the lower hind limb of hoofed animals, the lower portion of the leg of dinosaurs including birds or the rear paw. It is also represented in the rear 'paddle' of extinct marine reptiles, such as plesiosaurs. The oldest types of tetrapods had seven or eight digits.
 W
WSclerotic rings are rings of bone found in the eyes of many animals in several groups of vertebrates, except for extant mammals and crocodilians. They can be made up of single bones or multiple segments and take their name from the sclera. They are believed to have a role in supporting the eye, especially in animals whose eyes are not spherical, or which live underwater. Fossil sclerotic rings are known for a variety of extinct animals, including ichthyosaurs, pterosaurs, and dinosaurs, but are often not preserved.
 W
WA scute or scutum is a bony external plate or scale overlaid with horn, as on the shell of a turtle, the skin of crocodilians, and the feet of birds. The term is also used to describe the anterior portion of the mesonotum in insects as well as some arachnids.
 W
WThe synsacrum is a skeletal structure of birds and other dinosaurs, in which the sacrum is extended by incorporation of additional fused or partially fused caudal or lumbar vertebrae and it can only be seen in birds. Some posterior thoracic vertebrae, the lumbar, sacral and a few anterior caudal vertebrae are fused to form a complex bone called synsacrum. The innominate bones are fused with the synsacrum to a greater or lesser extent, according to species, forming an avian pelvis. This forms a more extensive rigid structure than the pelvis of a mammal, fulfilling requirements for flight, locomotion and respiration. Posterior to the synsacrum there are a few free caudal vertebrae, the last of which is the pygostyle to which the long, stiff tail feathers are attached. The central section of the synsacrum is swollen to accommodate the glycogen body, an organ whose function is as yet unclear but which may be associated with balance.
 W
WA thagomizer is the distinctive arrangement of four to ten spikes on the tails of stegosaurid dinosaurs. These spikes are believed to have been a defensive measure against predators.